
Microbiota of the weathering crust from the Turgoyakskoye kaolin deposit (Miass district, Southern Urals)
- 1 — Junior Researcher Peoples’ Friendship University of Russia named after Patrice Lumumba ▪ Orcid ▪ Elibrary
- 2 — Ph.D. Leading Researcher Borissiak Paleontological Institute of the RAS ▪ Orcid
- 3 — Ph.D., Dr.Sci. Professor Peoples’ Friendship University of Russia named after Patrice Lumumba ▪ Orcid
- 4 — Ph.D. Associate Professor Peoples’ Friendship University of Russia named after Patrice Lumumba ▪ Orcid
- 5 — Ph.D. Head of Department Peoples’ Friendship University of Russia named after Patrice Lumumba ▪ Orcid
Abstract
In recent decades, researchers have identified an important and in some cases leading role of biological processes, primarily microbiota activity, in the formation of weathering crusts. The study of new features associated with crust formation is of both theoretical and practical interest. The aim of this work is to clarify the role of microbiota in the formation of the Mesozoic kaolinite-type weathering crust (exemplified by the Turgoyakskoye kaolin deposit) using microprobe analysis and electron microscopy. The research showed active involvement of diverse microbial communities in the formation of kaolin clays: scientists found various fossilized bacterial structures, including mineralized biofilms, pseudomorphs after coccoid and rod‑shaped bacteria, glycocalyx remnants, and filamentous microfossils. This microbiota is morphologically similar to bacterial forms described in laterites, but the biominerals composing them are fundamentally different. In kaolins, they consist of silicates and aluminosilicates, while in laterites they consist of Fe, Al, and Mn oxides and hydroxides, which indicates a different species composition of microbiota involved in the formation of lateritic and kaolinitic weathering crusts. These findings likely show that during weathering, climate controls not only chemical but also bacterial mineral formation. Unlike laterites, the distribution of microorganisms in kaolin clays is uneven – ranging from isolated remnants to mass accumulations. During kaolin formation, microbiota biomechanically destroys the matrix of the parent rock and simultaneously synthesizes new minerals, forming biomorphs from them. Among the latter, biomorphs after “titanium” bacteria are of particular interest. Their discovery confirms Academician V.I.Vernadskii’s view on the predominantly biogenic nature of titanium in rocks of mature weathering crusts.
None
Introduction
One of the fundamental geological processes transforming our planet is the formation of sedimentary rocks, which is inextricably linked to weathering crusts of igneous, metamorphic, and ancient sedimentary complexes [1-3]. During their weathering, solid, liquid, and gaseous substances emerge; the accumulation of these substances gives rise not only to sedimentary rocks but also to a wide range of valuable minerals [4-6]. Over more than a century of studying weathering crusts, geologists have deciphered the mechanism of their formation [7-9]. This mechanism is based on deep chemical transformation of parent rocks in the hypergenesis zone under the influence of hydrolysis, hydration, oxidation, and carbonatization reactions. The resulting reaction products are utilized in the form of newly formed minerals and are also carried away by waters draining the weathering crusts. As a result, deposits accumulate on the parent rocks: at the bottom, they consist of clay minerals (lithomarge zone), and at the top – of oxides and hydroxides of Fe, Al, Mn, and Ti (laterite zone) [10-12]. The role of the biogenic factor in crust formation remains disputable. In the early 20th century, geologists such as V.I.Vernadskii, A.E.Fersman, B.B.Polynov, I.I.Ginzburg, as well as microbiologists V.L.Omelyanskii and S.N.Vinogradskii, wrote in their works about the possible active involvement of microorganisms in this process [13-15]. However, genuine interest in the microbial aspects of weathering crusts coincided with the period when the industry began to actively adopt biotechnological methods for extracting valuable chemical elements from ores using microorganisms. In the search for effective “working” microbial communities, researchers in various countries isolated strains of bacterial cultures aggressive towards minerals of different rock types [16-18]. For scientists, this circumstance became key to understanding weathering crusts and determined the direction of their further research.
Problem statement
For a long time, searches for biota in weathering crusts yielded no results, although laboratory experiments by microbiologists showed that prokaryotic microorganisms are capable of destroying rock-forming minerals, even complex ones such as silicates and aluminosilicates [19-21]. In nature, these processes can be observed on the surfaces of rocks colonized by bacterial‑algal biota, mosses, and lichens.
The situation changed radically when electron microscopy methods became ingrained in lithological research practice. Detailed study of rocks showed that, in addition to chemical processes, the activity of microbial organisms plays an important and in some cases dominant role in crust formation [22]. This is confirmed by hundreds of electron microscopic images showing fossilized filamentous, rod‑shaped, coccoid, and other bacterial remains. A surge in publications on identified microbiota occurred in the 1990s-2000s and was mainly associated with bauxites. The authors of a collective monograph provided a kind of guide to bacterial forms in bauxites, presenting biomorphic structures of bauxites from nearly fifty deposits across different regions of the world, with an age range from Late Proterozoic to Quaternary [23]. Using electron microscopy methods, geologists unexpectedly discovered a remarkable world of mineralized microorganisms, whose remains (biomorphs) turned out to be composed of mineral microparticles of Fe, Al, Mn, and Ti oxides and hydroxides. The idea of an inseparable link between biota and bauxites took a different tone at the current stage of research: groups of nanominerals were identified with proven microbial origin, which led to the emergence of a new term in specialized literature – “nanobiominerals” [24, 25]. Thus, thanks to electron microscopy methods, the active involvement of the biogenic factor in lateritic crust formation is beyond doubt. However, all of the above does not mean that weathering crusts are fully studied. On the contrary, many mysteries are still associated with them. Here are some of them:
- Where does the iron and manganese in laterites come from? The rocks of the kaolinite zone, on which laterites lie and which undergo lateritization, are characterized by negligible contents of these elements.
- What is the reason for known cases of direct contact between laterites and parent rocks?
- What is the nature of the so‑called “gingerbreads” – inclusions of unaltered parent rocks in lateritic bauxites?
- What is the nature of Ti concentrations in laterites and bauxites?
- What is the role of microbiota in kaolinitic weathering?
In this article, the authors will attempt to answer questions about the nature of Ti concentration in laterites and bauxites and the role of microbiota in kaolinitic weathering, as well as to determine the role of microorganisms in the formation of clay deposits in the lithomarge zone of weathering crusts, which is significantly less obvious than in the formation of laterites and bauxites. There are isolated general‑purpose publications on this issue [23, 26, 27], but the ability of soil biota to transform aluminosilicates into clay minerals is known for quite a long time [28-30]. This allows us to expect the existence of similar processes among the clays of the lithomarge. The relevance of the raised issue is obvious from both theoretical and practical points of view, since knowledge of all the details of crust formation significantly increases the efficiency of prospecting for minerals associated with weathering crusts. In this regard, the authors studied the lithomarge zone of the weathering crust at the Turgoyakskoye kaolin deposit in the Miass district of the Chelyabinsk Region, Southern Urals (Fig.1).
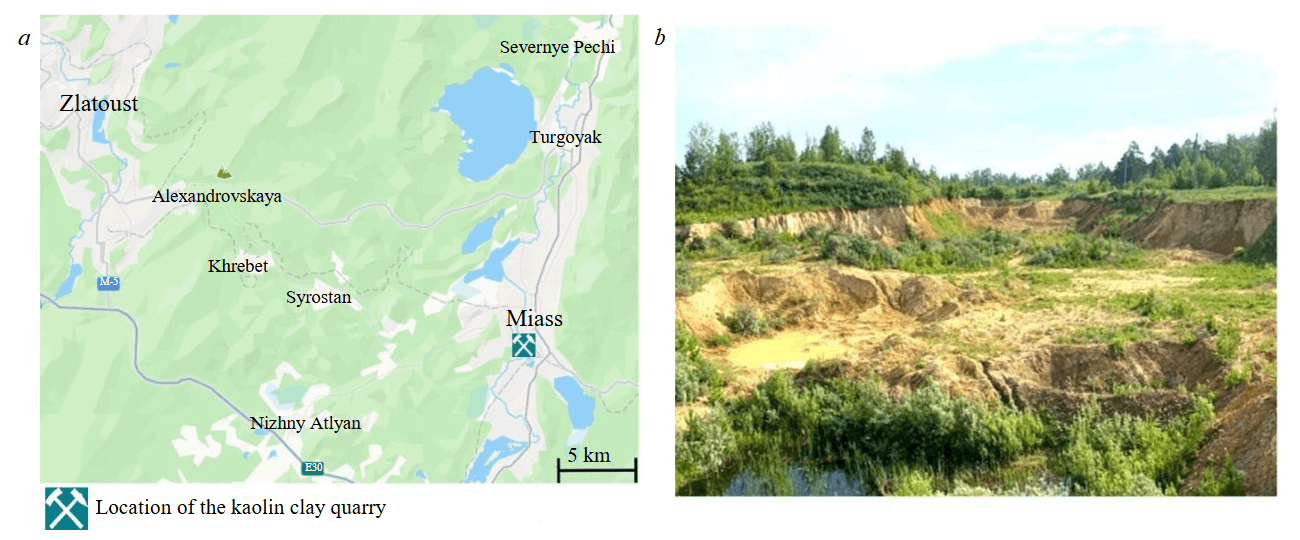
Fig.1. Location (a) and panorama (b) of the Turgoyakskoye kaolin deposit
The deposit is two hundred metres east of the active quarry for fluxing limestones and represents a tectonic block associated with the southeastern limb of a large anticlinal structure composed of the Lower Devonian rocks. Its core consists of fluxing limestones, while the limbs are formed by unevenly interbedded siliceous-clayey and carbonaceous-clayey shales. Since the early 21st century, the deposit is mothballed, but prior to that it was mined by open-pit method down to a depth of 10 m. The exposed kaolin in the lower part of the quarry is predominantly white or light grey, whereas in the upper bench it is variegated – brown, red, yellow, and crimson. A characteristic feature of the light-coloured kaolin is the widespread development of relicts of weathered shale rocks with preserved elements of a southeastern dip at an angle of 70-75° (Fig.2).
The presence of such relicts provides indisputable evidence that the kaolin clays belong to the eluvial type, which is genetically linked to the lithomarge zone of the weathering crust. The overlying sequence of the deposits in question is a near-horizontally bedded sand-clayey member with thin lenticles of lignite and quartz gravel at its base. Given the absence of fragments of ferruginous rocks in the gravel material, the observed weathering crust profile should evidently be classified as kaolinitic. The age of weathering is tentatively estimated as Mesozoic [3, 7], although the possibility of a Devonian stage of crust formation cannot be ruled out.
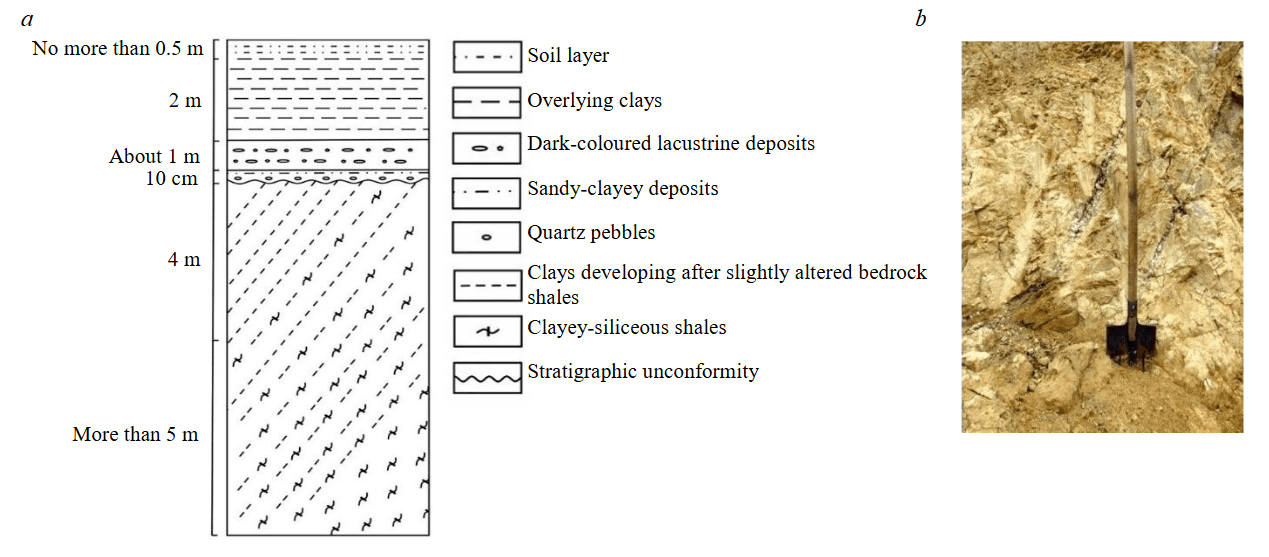
Fig.2. Lithological section (a) and a fragment of the kaolinite weathering crust from the lithomarge zone (b) of the Turgoyakskoye deposit
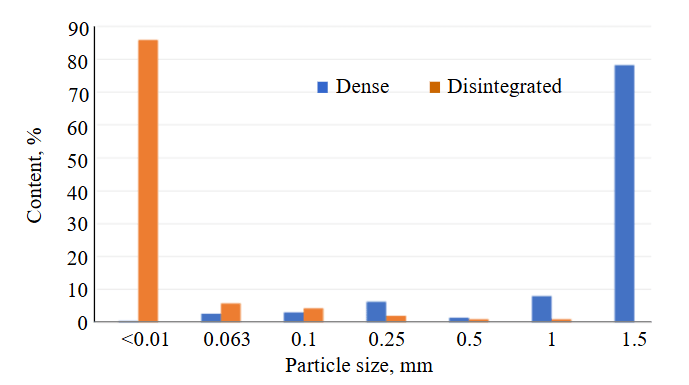
Fig.3. Results of particle size analysis of kaolin from the Turgoyakskoye deposit

Fig.4. Microscopic structure of kaolin.
Thin section XPL II. Visible field 2 mm
Materials and methods
The material for the article was provided by clay samples collected across section intervals and according to their varieties (colour range, degree of rock weathering, development of relict features of parent shales in the clays). The collected samples, in addition to microscopy (using a POLAM L213M polarizing microscope), underwent particle sizing sieve analysis; subsequently, the –0.01 mm fraction was studied using an Ultima-IV X-ray diffraction system (Rigaku, Japan). To examine the clastic material of larger fractions (+2-5 mm), a Tescan Vega 3 scanning electron microscope equipped with an INCA microanalysis system was used. Analytical work was carried out in the laboratories of Peoples’ Friendship University of Russia named after Patrice Lumumba (grain size measurements); Institute of Geology of Ore Deposits, Petrography, Mineralogy, and Geochemistry of the RAS (X-ray phase analysis); Borissiak Paleontological Institute of the RAS (electron microscopic studies).
Results
Lithological features of the clays. Documentation of the quarry walls showed that the degree of shale weathering is highly uneven and varies depending on the intensity of their schistosity. As a result, both along the section and along the strike, a chaotic alternation of intervals (1-10 cm) of relatively “dense” rocks and rocks transformed into fine grit and fine crumbs is observed. In turn, the so-called “dense” rocks easily crumble by hand and disperse in water to a clay-like state. The clear difference between these varieties is demonstrated by the results of grain size measurements, which includes settling clay particles and sieving of coarser material (Fig.3).
As the data show, the yield of –0.01 mm fraction should evidently be regarded as an indicator of the weathering activity and the intensity of clay mineral development after shales. In thin sections, weathered shales consist of a fine-to-microcrystalline aggregate of quartz-illite composition. Their structure preserves microcleavage, schistosity, as well as shadow outlines of leached or replaced primary minerals (Fig.4). The composition of the samples is specified by X-ray diffraction quantitative analysis data (see Table).
Mineral composition of clays from the weathering crust of the Turgoyakskoye deposit, %
|
Clay varieties |
Minerals and their main X-ray reflections |
|||||||||
|
Smectite (12.5-15.5А) |
Chlorite (14.2А) |
Kaolinite (7.15А) |
Micaceous (10.0А) |
Quartz (3.34А) |
Plagioclase (3.22А) |
Rutile (1.69А) |
Goethite (4.18А) |
Hematite (2.69А) |
|
|
|
“Dense” |
4.0 |
– |
8.1 |
43.8 |
44.1 |
– |
|
|||
|
Loose |
13.2 |
– |
26.6 |
26.8 |
29.9 |
– |
0.9 |
2.6 |
|
|
|
17.5 |
– |
16.1 |
30.1 |
15.0 |
21.3 |
– |
– |
– |
|
|
|
16.1 |
2.7 |
31.8 |
29.0 |
15.4 |
2.8 |
0.6 |
1.6 |
|
||
Based on clay minerals, loose (dispersed) clays of the weathering crust are more hypergenically altered than the “dense” varieties.
Results of studying clays from the weathering crust of the Turgoyakskoye deposit using scanning electron microscopy (SEM)
The studies were carried out using a Tescan Vega 3 instrument equipped with an INCA x-sight X-ray spectrometer with energy dispersion, 133 eV resolution and an active crystal area of 10 mm2. Sample chips no larger than 2×2 cm were coated with gold to improve electrical conductivity. Semi-quantitative chemical analysis was performed at an accelerating voltage of 20 kV and a current of 70 nA. The electron probe diameter was 0.3-3 µm, with an element detection range from 0.1 to 100 %. Sample examination was conducted using backscattered electrons (BSE) and secondary electrons (SE).
Given the physical state of the clays, insights into their structure are based on the study of “dense” rock samples, the features of which are documented by more than 200 electron microscopic images and microprobe measurements of rock‑forming minerals. Analysis of the collected material allows us to state that diverse microbial communities actively participated in the formation of kaolin clays at the Turgoyakskoye deposit. This is illustrated by numerous images of fossilized bacterial structures in the form of mineralized biofilms, pseudomorphs after colonies and single forms of coccoid and rod‑shaped bacteria, glycocalyx remnants, filamentous microfossils, as well as microbiota of unclear taxonomic position. The distribution of microorganisms in the clays is uneven – ranging from isolated remnants to mass accumulations. The composition of mineralized microbiota also varies widely, from oxide to silicate and aluminosilicate mineral varieties.
Characteristics of the found microbiota
Biofilms and their structureless fragments (glycocalyx) are the most common biomorphoses. They are observed on the walls of fractures and pores, on the surfaces of mineral grains, as well as on chips and on foliated planes of weathered rocks. It is assumed that such formations represented communities of microorganisms enclosed in a mucous substance made of polysaccharides (exopolysaccharide polymer substance, EPS), which are produced by bacterial cells for protection from external influences [31]. In the fossil state, the fossilization of such a substance results in mineral films with a bacterial or homogeneous (structureless) structure. From a morphological standpoint, the films are quite diverse and differ in appearance, degree of crystallization, and structural features. In the latter case, they can be either single- or multi-layered with alternating micro-cuts of micaceous particles. The surfaces of the films are smooth or finely bumpy. Their bumpy appearance is due to the contours of bacterial cells protruding from beneath the EPS cover (Fig.5, a). Details of the internal structure of the biofilms are revealed under high magnification. The main structure-forming forms are coccoid and rod-shaped bacteria. In multilayered films, the identified microfossils develop separately from one another, forming their own “environmental niche” within each layer.
Rod-shaped bacteria are elongated, cylindrical cells with rounded ends. The rods are straight, tightly pressed against each other, and therefore have a similar spatial orientation, which likely reflects the direction of movement of pore water, which is essential for the life of microorganisms. The rods are up to 25 µm long, and no more than 2.5 µm thick (Fig.5, b). Coccoid bacteria are tiny (0.5-1.0 µm) spherical bodies with clear, or less commonly, fuzzy, boundaries. By adhering to each other, they form massive clusters that fill depressions and overgrow the convex surfaces of biofilms (Fig.5, c). During crystallization of biofilms, the contours of coccoid cells are usually “erased,” but even in these cases, shadow relics of them remain (Fig.5, d). Mineralized disc-shaped bacteria develop separately from biofilms. They are typically found in groups of 10-15 individuals and differ from cocci not only in their distinctive shape but also in their significantly larger size, reaching 5 µm in diameter and 0.5 µm in thickness. The well-preserved biota allows us to observe cell division with their subsequent transformation into diplo- and tetracoccal forms, as well as sarcins (Fig.5, e). Similar bacterial remains were described in lateritic bauxites of Brazil and are attributed to cyanobacteria of the genus Synechococcus [23].

Fig.5. Secondary electron (SE) images of microbial structures in kaolin clays from the Turgoyakskoye deposit: a – fragment of mineralized biofilm (bf) with signs of replacement of a micaceous-montmorillonite aggregate (mn); the finely bumpy structure of the film is clearly visible, reflecting the contours of bacterial cells (bc) embedded in the film material; b – community of tightly pressed rod-shaped bacteria (r); c – massive clusters of coccoid microorganisms (c); d – recrystallized biofilm with shadow relics of bacterial cocci (rc) with an enlarged fragment in the inset; e – fossilized remains of discoid bacteria (s – sarcins, d – diplococci, t – tetracocci) and their energy-dispersive spectrum (f)
Mineralized microcolonies consist of biomorphs formed by spherical cells that, growing on top of each other, create grape-like structures on the walls of fractures (Fig.6, a). Such cells are characterized by clear boundaries, uniform size (8-10 µm), distinct division marks, a “wrinkled” surface, and an internal structure recrystallized into radial-fibrous patterns. Some of the spherical cells are flattened, which may indicate that this microbiota formed synthetically alongside rock compaction processes.
Biomorphs of filamentous microfossils are represented by remnants of cyanobacteria, actinomycetes, and micromycetes. This type of microbiota most often colonizes areas with clearly visible signs of mineral matrix destruction in the rock. A clear tendency for the noted microorganisms to be spatially separated from each other according to species-specific “environmental niches” is observed. Cyanobacterial remnants are mostly poorly preserved. They appear as single straight or curved flattened filamentous forms, in some cases retaining intact structure. In such cases, one can observe sheath fragments with a cross‑section of 1.5-2.5 µm and twisted trichomes enclosed within them (Fig.6, b). Mineralized actinomycetes resemble moss in electron images. This similarity is due to the dense interweaving of submicroscopic hyphae, which form a fuzzy coating on mineral rock particles (Fig.6, c). Myxomycetes consist of hyphae forming a mycelium of tangled, branching filaments 20-30 µm long. Many of them display thickenings 0.5-1 µm in size at their ends, which may represent fruiting bodies of microfungi. The preservation state of filamentous hyphae varies from well-defined forms to filaments whose outlines literally merge with the mineral matrix of the rock (Fig.6, d).
Biomorphs of microfossils of unclear taxonomic position are characterized by diverse morphology. These include regular and flattened spheres and ellipsoids, mushroom-like structures, as well as organic remnants in the form of cups and shells. Their sizes range from a few micrometres to several dozen micrometres. The surface is smooth or complicated by outgrowths consisting of tiny plates and spheres. Internally, the biomorphs can be either hollow or monolithic. Apparently, some of them represent remnants of mineralized eukaryotes (Fig.6, e, f).

Fig.6. Microbial structures in kaolin clays from the Turgoyakskoye deposit: a – mineralized grape-shaped microcolonies, with their radial-fibrous internal structure in the upper right corner; b – fragments of the remains of cyanobacteria sheaths (sh) within which trichomes (t) intertwined into “bundles” are visible; c – remains of mineralized actinomycetes (a), like moss, overgrowing and replacing the scales of micaceous minerals, with an enlarged fragment of a mica scale with traces of bacterial corrosion; d – mineralized hyphae (h) of myxomycetes with thickenings at the ends of the filaments (fruiting bodies? – (fr), in the right and central parts, several hyphae “merge” with particles of the rock matrix, indicating their identical mineral composition; e, f – examples of microfossil biomorphoses of unclear systematic position (possible remains of eukaryotes?). Images a, c, f – in secondary electrons (SE), b, d – in backscattered electrons (BSE); e – in backscattered and secondary electrons
Substance composition of microbiota biomorphoses
Analysis of microprobe data shows that mineralized biogenic remnants consist of a wide range of predominantly lithophilic and siderophilic elements. At least 20 different combinations of these elements were identified. Moreover, the chemical composition of the biota – regardless of the species identity of the microorganisms – is not constant but varies from oxide to silicate and aluminosilicate mineral varieties. Despite the apparent randomness in element distribution, two types of biomorphs emerge based on the pattern of their concentrations. The first type is compositionally close to or analogous to the surrounding mineral matrix of the rock (biofilms, coccoid communities, filamentous organisms). The second type clearly differs from it and represents isolated mineral phases (disc-shaped bacteria).
Biofilms exhibit the greatest compositional diversity. Their composition ranges from simple silicon oxide to complex multicomponent systems, which – in addition to Al, Fe, Mg, Ca, K, and Na – include trace amounts of Ti, Co, Sb, In, Tb, as well as Hf, Sc, and Er (Fig.7, a). In some cases, carbon is also present in this profile, which confirms the biogenic nature of the mineralized films. Due to the complex and variable set of constituent elements, it is not possible to determine the exact mineral composition of the films. However, necessary (albeit indirect) data were obtained by comparing microprobe analyses of the films and the mineral aggregates after which they develop. This comparison shows that in most cases these formations are chemically related and, therefore, should also be mineralogically similar. Thus, the films are determined to have kaolinitic, smectitic, mica-like, and siliceous compositions. As for the previously noted trace impurities, their presence evidently indicates the ability of biofilms to selectively accumulate elements whose concentrations in the mineral matrix of the rock are below the detection limit of microprobe analysis. This also applies to other types of microbiota, although they are clearly depleted in trace impurities. As an example, Fig.7, b shows siliceous biomorphs formed after remnants of micrococcal organisms that develop on the surface of a quartz grain.
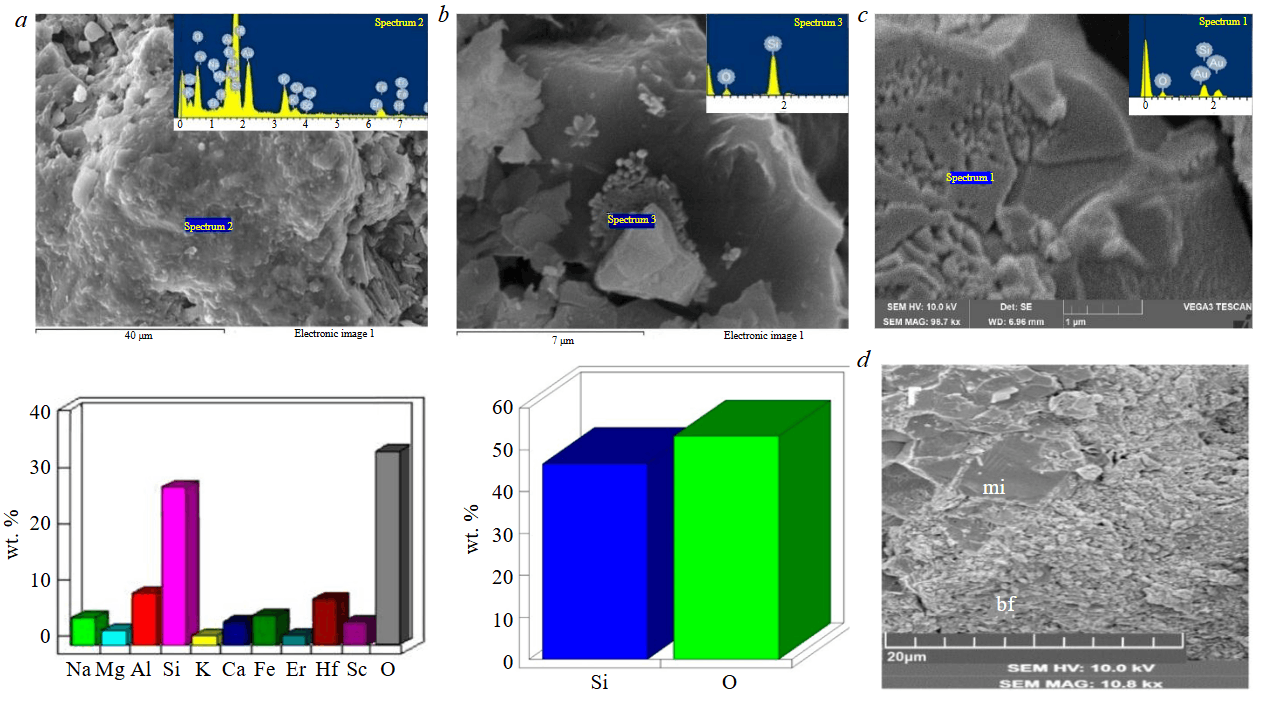
Fig.7. Secondary electron (SE) images of microbial structures in kaolin clays from the Turgoyakskoye deposit: a – biofilm fragment (glycocalyx) and its chemical composition (based on EDS data); b – biomorphoses after coccoid cells and their chemical composition (based on EDS data); c – traces of bacterial corrosion of a quartz grain; d – replacement of the micaceous aggregate (mi) of the rock matrix by biofilm material;
relict mica particles are clearly visible on the biofilm surface
A very important question is in what form does the substance composing the noted biomorphs exist? The observations show that both crystalline and amorphous phases are involved in their structure. In some cases (for example, for biofilms), a combined state of the substance is recorded, with transitions from structureless X-ray-amorphous (?) areas to weakly and well-crystallized sections. In other words, a clear transformation of the substance during its recrystallization – from the amorphous to the crystalline state – can be traced. This is important for understanding the formation mechanism of the previously noted radial-fibrous structures of bacterial colonies. Obviously, they also result from crystallization of smectite mineral metacolloids.
A different pattern is characteristic of disc‑shaped bacteria. The composition of their biomorphs, regardless of surrounding minerals, is always uniform and represented by titanium dioxide (see Fig.5, f), sometimes enriched with W and Rb. Here, a clear in-life specialization of the biota with respect to titanium emerges. As in other cases, it is not possible to accurately determine the mineral identity of disc-shaped biomorphs, since 11 mineral modifications of titanium dioxide are known, the most common of which are rutile, brookite, and anatase [32].
The role of microbiota in the formation of the weathering crust of the Turgoyakskoye deposit
The identified morphological, mineralogical, and compositional characteristics of microbiota, as well as the features of its relationship with rock-forming minerals, allow us to assess the contribution of microorganisms to kaolinitic crust formation. To understand the problem, it is advisable to compare the presented material with known data on the role of microbiota in the formation of lateritic weathering crusts. Studies by domestic and foreign scientists found the widespread and multifaceted involvement of bacterial communities – filamentous, coccoid, and rod-shaped organisms – in lateritic weathering, whose activity has both destructive and constructive aspects [23, 25, 26]. The destructive aspect manifests as mechanical destruction and biochemical leaching of mineral aggregates, which leads to dispersion of weathered rocks. The constructive aspect appears in the accumulation of dissolved matter and its deposition in the form of pseudomorphs after biofilms, remnants of cyanobacteria, and filamentous coccoid and rod-shaped microfossils. This results in the formation of metacolloidal, dispersed, and crystallized mineral phases of oxides and hydroxides of Al, Fe, Mn, and Ti.
Comparative analysis of the presented materials on microbiota in laterites and kaolins from the Turgoyakskoye deposit allows us to identify similarities and differences in biogenic processes of the two types of crust formation. In particular, the same bacterial morphotypes are actively involved in kaolinitic weathering mechanisms as in laterites, but their density is noticeably lower, and the microorganisms themselves are predominantly isolated in different environmental niches. Similar to laterites, multifaceted activity of microbiota is also observed in kaolin rocks. On the one hand, this activity leads to destruction of the mineral matrix of rocks; on the other hand, it contributes to its restoration through synthesis of new (“biogenic”) minerals. Bacterial destruction of the matrix is achieved through mechanical and biochemical effects of microorganisms on minerals. Most often, these two factors manifest jointly, complementing each other. The mechanical factor is fully evident in filamentous organisms when their trichomes or hyphae, growing, penetrate between mineral flakes and push them apart (see Fig.6, d). At the same time, bacterial cells, by releasing metabolites, chemically dissolve surrounding minerals and thereby subject the kaolin rock to additional dispersion. As observations show, traces of biochemical dissolution can be found even on the surface of quartz grains (Fig.7, c).
Bacteria and bacterial communities affect minerals not only indirectly but also directly, causing their corrosion. In this case, replacement structures form along the contacts (see Fig.6, c), with biogenic tissue penetrating into mineral grains, leaving only relict particles behind (see Fig.5, a, 7, d). A certain relationship is observed between the biota and the composition of the replaced mineral aggregate. Micaceous minerals and feldspars are most actively subjected to microbial “attack”, followed by montmorillonite to a lesser extent, and kaolinite is affected extremely rarely. Potassium may be the possible cause of this phenomenon. On the one hand, it is part of the mineral composition; on the other, it is a chemical element essential for bacterial life. Probably, this is the reason why bacteria primarily corrode minerals most enriched in potassium – especially when it is in an easily accessible state. The possible course of events was found through experimental work and geological observations [16, 20, 33].
Another important issue concerns biogenic minerals in the kaolinitic weathering crust. Like in laterites, they are an important component of the forming rocks and are also represented by pseudomorphs after biogenic remnants. The general mechanism of their formation is discussed in a series of works by Russian and foreign scientists [31, 34, 35]. This process is divided into biologically induced mineralization (BIM) and biologically controlled mineralization (BCM) [36-38]. In the first case, minerals crystallize on cell surfaces and their membranes through reactions between metabolic products and mineralized pore solutions. In the second case, mineral nucleation and growth occur inside cells and are controlled by intracellular processes. Despite the same biota composition and identical mechanism of biogenic mineral formation, the results of microbial activity under lateritic and kaolinitic weathering conditions differ fundamentally. This is clearly seen in the composition of authigenic biominerals: in laterites, they are represented by oxides and hydroxides of Fe, Al, Mn, and Ti; in kaolins, they consist of silicate and aluminosilicate varieties, among which clay biominerals of colloidal size strongly dominate. In both types of weathering crusts, the composition of biominerals is similar to abiogenic minerals formed chemically during rock weathering. This leads to the conclusion that during weathering crust formation – regardless of its type – chemical and biogenic mineral formation follow the same laws, which are controlled by the climatic factor and directly depend on the composition of parent rocks. As is known, these two factors determine the activity of the geochemical environment – its Eh, pH, temperature, and the intensity of water drainage in weathered rocks. In turn, the geochemical conditions of weathering (alkaline for kaolins and acidic for laterites) determine the ability of elements to transfer into solutions and participate in chemical and biological reactions. In laterites, judging by the ratio of chemogenic and biogenic ultramicrostructures, biogenic mineral formation processes successfully compete with chemical ones [24, 25, 39].
In the kaolins of the Turgoyakskoye deposit, the role of biota is not so obvious. Its influence is most pronounced in local fractured zones, where biogenic structures and, consequently, biominerals become dominant in the rock texture. Here, another important point should be noted. In both types of weathering crusts under consideration, remnants of microorganisms of the same appearance are recorded (cocci, bacilli, filamentous forms), but the minerals they form are sharply different: oxides and hydroxides in one case, silicates and aluminosilicates in the other. This leads to the conclusion that the species composition of microorganisms involved in lateritic and kaolinitic crust formation differs. It can be concluded that in kaolinitic crusts, microbiota, by forming biominerals, acts as a kind of intermediate collector where dissolved substances from mineral aggregates destroyed by weathering are temporarily accumulated. Subsequently, either biominerals are conserved for an indefinite period, or – when weathering transitions to the lateritic stage – they are transformed by chemical and microbial processes into oxides and hydroxides of Fe, Al, Mn, and Ti.
A special place among the microbiota is occupied by “titanium” bacteria capable of accumulating TiO2 (see Fig.5, e). The appearance of titanium biomorphs in kaolin rocks contradicts the geochemical characteristics of titanium, which is practically immobile under hypergenic conditions. Its concentrations in laterites and bauxites are usually explained by the accumulation of residual titanium mineral grains that survived after weathering of the parent rocks [40-42]. However, as early as in the 1930s, V.I.Vernadskii, based on evidence of Ti concentration in organisms, proposed the existence of bacteria in nature capable of assimilating this element [15]. It is their activity that may be the main cause of titanium concentration in the end products of weathering (laterites and bauxites). The results of electron microscopic studies of kaolins from the Turgoyakskoye deposit confirm the correctness of the eminent scientist’s hypothesis.
Conclusion
The conducted studies show that the Turgoyakskoye deposit kaolin represents a kaolinitic weathering crust developed after shales. In addition to kaolinite itself, its composition includes micaceous and smectite minerals, as well as particles of feldspars and quartz processed to varying degrees. A crucial structural element of the kaolin is biomorphs after bacterial remnants of various morphologies (cocci, bacilli, bacterial films, filamentous organisms, glycocalyx). Unlike laterites, their mass accumulation is limited to local zones – especially in areas where the rock shows clear signs of degradation (porosity, fracturing).
Observations indicate that during kaolin formation, microbiota participate in two processes: they biomechanically destroy the matrix of the original rock and simultaneously synthesize new minerals, forming biomorphs. Biota mineralization is accompanied by precise replication of the morphological features of bacterial cells. This may serve as evidence of its belonging to the biologically induced type [36, 38]. It is also fundamental that, despite the analogy in morphological types of bacterial remnants in laterites and kaolins, the composition of biominerals in biomorphs is completely different: in laterites, these are oxides and hydroxides of Fe, Al, and Mn; in kaolins, these are silicates and aluminosilicates. This directly indicates a different species composition of microbiota involved in the formation of lateritic and kaolinitic weathering crusts. From this, another conclusion emerges: since the main factor of crust formation is climate, during weathering it controls not only chemical but also bacterial mineral formation.
Traditionally, titanium accumulations are explained by concentrations of residual titanium minerals preserved from the weathering of parent rocks [40-42]. However, Academician V.I.Vernadskii predicted a possible bacterial mechanism for element accumulation in laterites and bauxites [15]. Microprobe and electron microscopic studies of the Turgoyakskoye kaolin deposit revealed remnants of disc‑shaped titanium-specialized bacteria capable of accumulating titanium by extracting it from surrounding minerals. This fact, on the one hand, confirms V.I.Vernadskii’s view, and on the other, expands our understanding of titanium’s nature in weathering crusts, where it exists not only in autochthonous minerals but also in biogenic form. Assessing the ratio of these titanium varieties is a task for future research.
References
- Polynov B.B. Weathering crust. Part 1. Weathering processes. Main phases and forms of the weathering crust and their distribution. L.: Izd-vo Akademii nauk SSSR, 1934, p. 243.
- Petrov V.P. Fundamentals of the theory of ancient weathering crusts. Moscow: Nedra, 1967, p. 343.
- Ginzburg I.N. Ancient weathering crust on ultramafic rocks of the Urals. In 2 parts. Part 2. Geochemistry and geology of the ancient weathering crust in the Urals. Trudy Instituta geologicheskikh nauk. Moscow: Izd-vo Akademii nauk SSSR, 1947, p. 135.
- Slukin A.D. Weathering crusts and bauxites of ancient Asian platforms: Avtoref. dis. … d-ra geol.-mineral. nauk. Moscow: Institut geologii rudnykh mestorozhdenii, petrografii, mineralogii i geokhimii, 1991, p. 50.
- Dobrovolsky V.V. Hypergenesis and crusts of weathering. Selected Works. Vol. 1. Moscow: Scientific World, 2007, p. 512 (in Russian).
- Bowell R.J., Butt C.R.M. Geology, Geochemistry and Formation of Supergene Mineral Deposits in Deeply Weathered Terrain. Springer, 2025, p. 428. DOI: 10.1007/978-3-031-75733-4
- Razumova V.N., Kheraskov N.P., Chernyakhovskii A.G. Geological types of weathering crusts and examples of their distribution in the Southern Urals. Moscow: Izd-vo Akademii nauk SSSR, 1963, p. 142.
- Alcalde-Aparicio S., Vidal-Bardán M., Alonso-Herrero E. Mineralogy and geochemical signatures as indicators of differential weathering in natural soil profiles from the West Asturian-Leonese Zone (NW Iberia). Earth Sciences Research Journal. 2022. Vol. 26. N 1, p. 55-66. DOI: 10.15446/esrj.v26n1.81087
- Perri F. Chemical weathering of crystalline rocks in contrasting climatic conditions using geochemical proxies: in contrasting climatic conditions using geochemical proxies: an-overview. Palaeogeography, Palaeoclimatology, Palaeoecology. 2020. Vol. 556. N 109873. DOI: 10.1016/j.palaeo.2020.109873
- Mamedov V.I., Makarova M.A., Zaitsev V.A. The World’s Largest Fouta Djallon–Mandingo Bauxite Province (West Africa): Part III. The Influence of Geomorphic Factors and the Age of Relief on the Spatial Distribution, Size, and Quality of Bauxite Deposits. Geology of Ore Deposits. 2022. Vol. 64. Suppl. 1, p. 15-42. DOI: 10.1134/S1075701522090057
- Kai Deng, Shouye Yang, Yulong Guo. A global temperature control of silicate weathering intensity. Nature Communications. 2022. Vol. 13. N 1781. DOI: 10.1038/s41467-022-29415-0
- Gunathilake B.М., Jayawardana D.T., Ratnayake A.S. et al. Gondwana sedimentary rocks of Andigama Basin, Sri Lanka: unraveling weathering dynamics, tectonic setting, and paleoclimate. International Journal of Earth Sciences. 2024. Vol. 113. Iss. 8, p. 2357-2372. DOI: 10.1007/s00531-024-02423-9
- Polynov B.B. On the geological role of organisms. Voprosy geografii. Moscow: Gosudarstvennoe izd-vo geograficheskoi literatury, 1953. Sbornik 33. Fizicheskaya geografiya, p. 45-64.
- Fersman A.E. Selected Works. V 7 t. T. 5. Moscow: Izd-vo Akademii nauk SSSR, 1959, p. 858.
- Vernadskii V.I. Selected Works. V 5 tomakh. T. 5. Biosfera I-II. Stati po biogeokhimii. Pochvy. Gazy. Meteority i kosmicheskaya pyl. Moscow: Izd-vo Akademii nauk SSSR, 1960, p. 422.
- Štyriaková I., Štyriak I., Nandakumar M.P., Mattiasson B. Bacterial destruction of mica during bioleaching of kaolin and quartz sandsby Bacillus cereus. World Journal of Microbiology and Biotechnology. 2003. Vol. 19. Iss. 6, p. 583-590. DOI: 10.1023/A:1025176210705
- Sedlakova-Kadukova J., Marcincakova R., Luptakova A. et al. Comparison of three different bioleaching systems for Li recovery from lepidolite. Scientific Reports. 2020. Vol. 10. N 14594. DOI: 10.1038/s41598-020-71596-5
- Schmitz A.M., Pian B., Medinet S. et al. Generation of a Gluconobacter oxydans knockout collection for improved extraction of rare earth elements. Nature Communications. 2021. Vol. 12. N 6693. DOI: 10.1038/s41467-021-27047-4
- Avakian Z.A., Karavaiko G.I., Melnikova E.O. et al. Role of microscopic fungi in the process of weathering of pegmatite deposit rocks and minerals. Mikrobiologiia. 1981. Vol. 50. N 1, p. 156-162 (in Russian).
- Karavaiko G.I. Microbial destruction of silicate minerals. Trudy Instituta mikrobiologii im. S.N.Vinogradskogo: Yubileinyi sbornik k 70-letiyu instituta. Vyp. XII. Moscow: Nauka, 2004, p. 172-196.
- Song W., Ogawa N., Oguchi C.T. et al. Effect of Bacillus subtilis on granite weathering: A laboratory experiment. Catena. 2007. Vol. 70. Iss. 3, p. 275-281. DOI: 10.1016/j.catena.2006.09.003
- Antoshkina A.I. Bacterial rock formation is a reality of modern research methods. Uchenye zapiski Kazanskogo universiteta. Seriya Estestvennye nauki. 2011. Vol. 153. Book 4, p. 114-126 (in Russian).
- Shkolnik E.L., Zhegallo E.A., Bogatyrev B.A. et al. Biomorphic structures in bauxites (based on the results of electron microscopic study). Moscow: Eslan, 2004, p. 112.
- Slukin A.D., Boeva N.M., Zhegallo E.A., Zaitseva L.V. Laterite bauxite biominerals – new data from electron microscopic studies. New Data on Minerals. 2016. Vol. 51, p. 52-61 (in Russian).
- Novikov V.M., Bortnikov N.S., Boeva N.M. et al. Biogenic nanominerals of iron oxides from basalt weathering core of continental suburbs of East Asia on the example of the Far East of Russia and Vietnam. Paper 3. Magnetite. Proceedings of Voronezh State University. Series: Geology. 2017. N 2, p. 69-73 (in Russian).
- Xiaoxue Yang, Yanzhang Li, Yan Li et al. Microbially induced clay weathering: Smectite-to-kaolinite transformation. American Mineralogist. 2023. Vol. 108. N 10, p. 1940-1947. DOI: 10.2138/am-2022-8442
- Kawano M., Tomita K. Microbiotic Formation of Silicate Minerals in the Weathering Environment of a Pyroclastic Deposit. Clays and Clay Minerals. 2002. Vol. 50. Iss. 1, p. 99-110. DOI: 10.1346/000986002761002865
- Naimark E.B., Erouschev-Shack V.A., Chizhikova N.P., Kompantseva E.I. Interaction of clay minerals with microorganisms: a review of experimental data. Zhurnal obshchei biologii. 2009. Vol. 70. N 2, p. 155-167 (in Russian).
- Perelomov L.V. The Role of Interactions between Bacteria and Clay Minerals in Pedochemical Processes. Geochemistry International. 2023. Vol. 61. N 10, p. 1026-1035. DOI: 10.1134/S0016702923100105
- Sokolova T. The role of soil biota in the weathering of minerals: A review of literature. Eurasian Soil Science. 2011. Vol. 44. N 1, p. 56-72. DOI: 10.1134/S1064229311010121
- Astafeva M.M., Gerasimenko L.M., Geptner A.R. et al. Fossil bacteria and other microorganisms in terrestrial rocks and astromaterials. Moscow: Paleontologicheskii institut im. A.A.Borisyaka RAN, 2011, p. 172.
- Tkhant Zin Pyu. Development of a targeted method for obtaining photoactive inorganic coatings based on titanium dioxide modified with copper: Avtoref. dis. … kand. khim. nauk. Moscow: Rossiiskii khimiko-tekhnologicheskii universitet imeni D.I.Mendeleeva, 2023, p. 16.
- Štyriaková I., Štyriak I., Oberhänsli H. Rock weathering by indigenous heterotrophic bacteria of Bacillus spp. at different temperature: a laboratory experiment. Mineralogy and Petrology. 2012. Vol. 105. Iss. 3-4, p. 135-144. DOI: 10.1007/s00710-012-0201-2
- Tikhomirova N.S., Orleansky V.K. Modelling of phosphate sedimentation in laboratory cyanobacteria. Lithology and Mineral Resources. 1994. N 1, p. 135-140 (in Russian).
- Weiner S., Dove P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. N 1, p. 1-29. DOI: 10.2113/0540001
- Lowenstam H.A., Weiner S. On Biomineralization. Oxford University Press, 1989, p. 333. DOI: 10.1093/oso/9780195049770.001.0001
- Bazylinski D.A., Frankel R.B. Biologically Controlled Mineralization in Prokaryotes. Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. N 1, p. 217-247. DOI: 10.2113/0540217
- Frankel R.B., Bazylinski D.A. Biologically Induced Mineralization by Bacteria. Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. N 1, p. 95-114. DOI: 10.2113/0540095
- Xiluo Hao, Kwunlun Leung, Rucheng Wang et al. The geomicrobiology of bauxite deposits. Geoscience Frontiers. 2010. Vol. 1. Iss. 1, p. 81-89. DOI: 10.1016/j.gsf.2010.06.001
- Yudovich Ya.E., Ketris M.P., Rybina N.V. Geochemistry of titanium. Syktyvkar: Geoprint, 2018, p. 431.
- Strakhov N.M. On the significance of titanium modulus in the establishment of the genesis of bauxites. Lithology and Mineral Resources. 1963. N 2, p. 249-252 (in Russian).
- Bushinskii G.I. Geology of bauxites. Moscow: Nedra, 1975, p. 411.