
Микробиота коры выветривания Тургоякского месторождения каолина (Миасский район, Южный Урал)
- 1 — младший научный сотрудник Российский университет дружбы народов имени Патриса Лумумбы ▪ Orcid ▪ Elibrary
- 2 — канд. геол.-минерал. наук ведущий научный сотрудник Палеонтологический институт им. А.А.Борисяка РАН ▪ Orcid
- 3 — д-р геол.-минерал. наук профессор Российский университет дружбы народов имени Патриса Лумумбы ▪ Orcid
- 4 — канд. геол.-минерал. наук доцент Российский университет дружбы народов имени Патриса Лумумбы ▪ Orcid
- 5 — канд. геол.-минерал. наук заведующий кафедрой Российский университет дружбы народов имени Патриса Лумумбы ▪ Orcid
Аннотация
В последние десятилетия выявлена важная, а в ряде случаев ведущая роль биологических процессов и прежде всего деятельности микробиоты в образовании кор выветривания. Изучение новых объектов, связанных с корообразованием, представляет не только теоретический, но и практический интерес. Цель данной работы – выяснить роль микробиоты в формировании мезозойской коры выветривания каолинитового типа (на примере Тургоякского месторождения каолина) с использованием микрозондового анализа и электронной микроскопии. В результате проведенных исследований установлено активное участие в образовании каолиновых глин разнообразных сообществ микроорганизмов – выявлены различные фоссилизированные бактериальные структуры: минерализованные биопленки, псевдоморфозы по коккоидным и палочковидным бактериям, остатки гликокаликса, нитчатые микрофоссилии. Указанная микробиота морфологически аналогична бактериальным формам, описанным в латеритах, но слагающие их биоминералы принципиально иные. В каолинах они представлены силикатами и алюмосиликатами, а в латеритах – оксидами и гидроксидами Fe, Al, Mn, что говорит о разном видовом составе микробиоты, участвующей в формировании латеритных и каолинитовых кор выветривания. Это, вероятно, показывает, что при выветривании климат контролирует не только химическое, но и бактериальное минералообразование. В отличие от латеритов, распределение микроорганизмов в каолиновых глинах неравномерное – от единичных остатков до массовых скоплений. При создании каолина микробиота биомеханически разрушает матрицу исходной породы и одновременно с этим синтезирует новые минералы, формируя из них биоморфозы. Среди последних особый интерес вызывают биоморфозы по «титановым» бактериям. Их обнаружение подтверждает точку зрения академика В.И.Вернадского о преимущественно биогенной природе титана в породах зрелых кор выветривания.
Отсутствует
Введение
Одним из фундаментальных геологических процессов, преобразующих нашу планету, является образование осадочных горных пород, неразрывно связанное с корами выветривания магматических, метаморфических и древних осадочных комплексов [1-3]. В ходе их выветривания возникают твердые, жидкие и газообразные вещества, при аккумуляции которых рождаются не только осадочные породы, но и широкий спектр ценных полезных ископаемых [4-6]. За более чем столетний период изучения кор выветривания геологам удалось расшифровать механизм их образования [7-9]. В его основе лежит глубокая химическая переработка материнских пород в зоне гипергенеза под воздействием реакций гидролиза, гидратации, окисления и карбонатизации. Возникающие продукты реакций утилизируются в форме новообразованных минералов, а также выносятся водами, дренирующими коры выветривания. В результате на материнских породах накапливаются отложения, внизу сложенные глинистыми минералами (зона литомаржа), а вверху – минералами оксидов и гидроксидов Fe, Al, Mn, Ti (зона латеритов) [10-12]. Дискуссионным остается вопрос о роли биогенного фактора в корообразовании. О возможном активном участии микроорганизмов в этом процессе в начале прошлого века в своих трудах писали такие геологи, как В.И.Вернадский, А.Е.Ферсман, Б.Б.Полынов, И.И.Гинзбург и микробиологи В.Л.Омелянский и С.Н.Виноградский [13-15]. Однако подлинный интерес к микробной проблематике кор выветривания совпал с периодом, когда в промышленности стали активно внедряться биотехнологические методы извлечения из руд ценных химических элементов с помощью микроорганизмов. В поисках эффективных «рабочих» микробных сообществ в разных странах были выделены штаммы бактериальных культур, агрессивных к минералам различных горных пород [16-18]. Для ученых это обстоятельство стало ключевым в понимании кор выветривания и определило направление их дальнейших исследований.
Постановка проблемы
Долгое время поиски биоты в корах выветривания не давали результатов, хотя лабораторные эксперименты микробиологов показывали, что прокариотные микроорганизмы способны разрушать породообразующие минералы, даже такие сложноустроенные, как силикаты и алюмосиликаты [19-21]. В природе эти процессы можно наблюдать на поверхности скал, заселенных бактериально-водорослевой биотой, мхами и лишайниками.
Кардинально положение дел изменилось, когда в практику литологических исследований прочно вошли методы электронной микроскопии. Детальное изучение пород показало, что помимо химических процессов важную, а местами и главную роль в корообразовании играет деятельность микробных организмов [22]. Это подтверждают сотни электронно-микроскопических снимков с изображением фоссилизированных нитчатых, палочковидных, коккоидных и других бактериальных остатков. Всплеск публикаций по выявленной микробиоте приходится на 1990-2000-е гг. и связан он главным образом с бокситами. Своеобразный путеводитель по бактериальным формам в бокситах получился у авторов коллективной монографии, где приведены биоморфные структуры бокситов почти полусотни месторождений из разных регионов мира и возрастного интервала от позднего протерозоя до четвертичного периода [23]. Применяя методы электронной микроскопии, геологи неожиданно для себя увидели удивительный мир минерализованных микроорганизмов, остатки которых (биоморфозы) оказались сложенными минеральными микрочастицами оксидов и гидроксидов Fe, Al, Mn, Ti. Идея о неразрывной связи биоты с бокситами получила новое звучание на современном этапе исследований – выделены группы наноминералов, для которых доказано микробиальное происхождение, благодаря чему в специализированной литературе появился новый термин «нанобиоминералы» [24, 25]. Таким образом, благодаря методам электронной микроскопии активное участие биогенного фактора в латеритном корообразовании не вызывает сомнения. Вместе с тем, все сказанное не означает, что коры выветривания до конца изучены. Напротив, с ними связано еще много загадок. Вот некоторые из них:
- Откуда в латеритах железо и марганец? Породы каолинитовой зоны, на которых латериты залегают и которые подвергаются латеритизации, характеризуются мизерными содержаниями этих элементов.
- В чем причина известных случаев прямого контакта латеритов с материнскими породами?
- Какова природа так называемых «пряников» – включений неизмененных материнских пород в латеритных бокситах?
- Какова природа концентраций Тi в латеритах и бокситах?
- Какова роль микробиоты в каолинитовом выветривании?
В данной статье авторы постараются ответить на вопросы о природе концентрации Ti в латеритах и бокситах и роли микробиоты в каолинитовом выветривании, а также установить роль микроорганизмов при формировании глинистых отложений зоны литомаржа кор выветривания, которая значительно менее очевидна, чем при формировании латеритов и бокситов. По этому вопросу имеются единичные публикации общего плана [23, 26, 27], однако о способности почвенной биоты трансформировать алюмосиликаты в глинистые минералы известно достаточно давно [28-30]. Это позволяет надеяться на существование аналогичных процессов и среди глин литомаржа. Актуальность поднятого вопроса очевидна как с теоретической, так и практической точек зрения, поскольку знание всех тонкостей корообразования значительно повышает эффективность поисков полезных ископаемых, связанных с корами выветривания. В связи с этим авторами проведено изучение зоны литомаржа коры выветривания Тургоякского месторождения каолина в Миасском районе Челябинской обл. Южного Урала (рис.1).
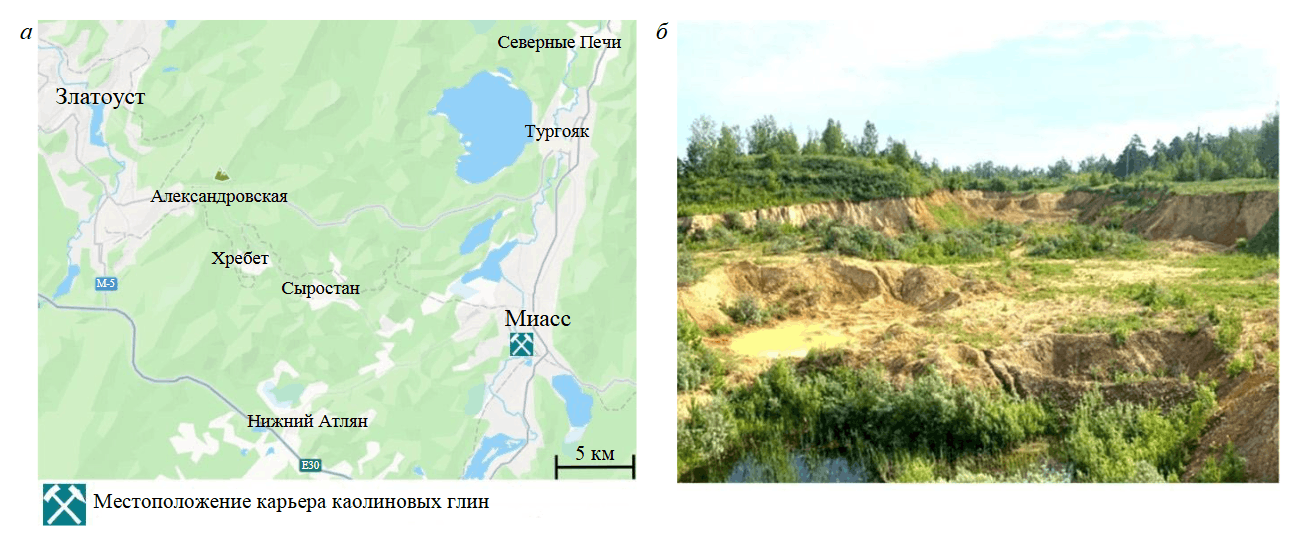
Рис.1. Местоположение (а) и панорама (б) Тургоякского месторождения каолина
Месторождение расположено в двухстах метрах восточнее действующего карьера флюсовых известняков и представляет собой тектонический блок, приуроченный к юго-восточному крылу крупной антиклинальной структуры, представленной породами нижнего девона. Ее ядро сложено флюсовыми известняками, а крылья – неравномерно переслаивающимися кремнисто-глинистыми и углеродисто-глинистыми сланцами. С начала XXI в. месторождение законсервировано, но до этого момента оно отрабатывалось открытым способом до глубины 10 м. Вскрытый каолин в нижней части карьера преимущественно белого или светло-серого цвета, тогда как в верхнем уступе он пестрый – бурый, красный, желтый, малиновый. Характерной чертой светлоокрашенного каолина является широкое развитие реликтов выветрелых сланцевых пород с сохранившимися элементами падения на юго-восток под углом 70-75° (рис.2).
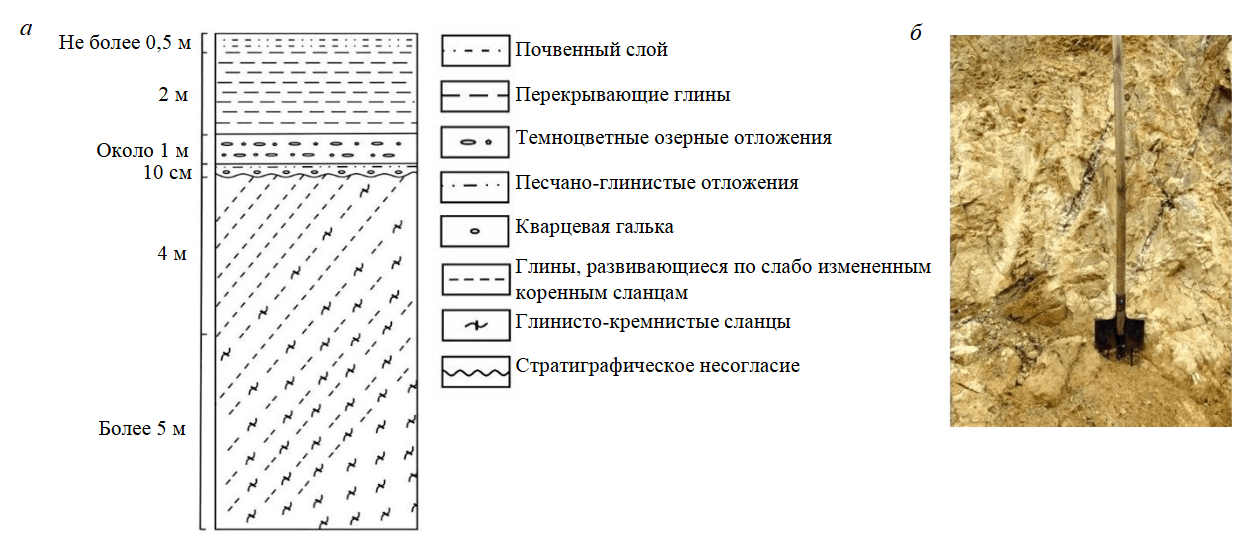
Рис.2. Литологический разрез (а) и фрагмент каолинитовой коры выветривания зоны литомаржа (б) Тургоякского месторождения
Присутствие таких реликтов служит бесспорным доказательством принадлежности каолиновых глин к элювиальному типу, который генетически связан с зоной литомаржа коры выветривания. Перекрывающей толщей рассматриваемых отложений служит субгоризонтально залегающая песчано-глинистая пачка с тонкими линзочками лигнита и кварцевого гравия в ее основании. Учитывая отсутствие в составе гравийного материала обломков ожелезненных пород, наблюдаемый профиль коры выветривания, очевидно, следует относить к каолинитовому типу. Возраст выветривания ориентировочно оценивается как мезозойский [3, 7], хотя нельзя исключать возможность существования и девонского этапа корообразования.
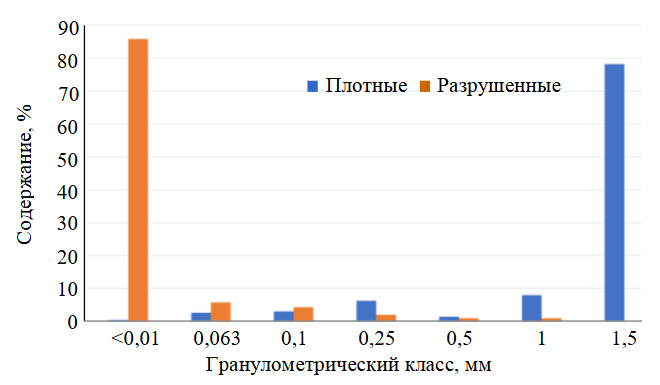
Рис.3. Результаты гранулометрического анализа каолина
Тургоякского месторождения

Рис.4. Микроскопическое строение каолина. Шлиф. Ник. II. Видимое поле 2 мм
Материалы и методы исследований
Материалом для статьи послужили образцы глин, отобранные по интервалам разреза, а также согласно их разновидностям (цветовой гамме, степени выветренности пород, развитию в глинах реликтовых признаков материнских сланцев). Отобранные пробы, помимо микроскопии (поляризационный микроскоп ПОЛАМ Л213М), подвергались гранулометрическому рассеву, после чего класс –0,01 мм изучался при помощи рентгеновского дифрактометра Ultima-IV фирмы Rigaku (Япония). Для исследования обломочного материала крупных классов (+2-5 мм) использовался сканирующий электронный микроскоп Tescan Vega 3, оснащенный микроанализатором системы INCA. Аналитические работы проводились в лабораториях Российского университета дружбы народов имени Патриса Лумумбы (гранулометрический анализ), Института геологии рудных месторождений, петрографии, минералогии и геохимии РАН (рентгенофазовый анализ) и Палеонтологического института имени А.А.Борисяка РАН (электронно-микроскопические исследования).
Результаты исследований
Литологические особенности глин
Документация бортов карьера показала, что степень выветривания сланцев крайне неравномерна и меняется в зависимости от интенсивности их рассланцевания. Благодаря этому, как по разрезу, так и по простиранию наблюдается хаотичное чередование интервалов (1-10 см) относительно «плотных» пород и пород, превращенных в мелкую дресву и тонкую крошку. В свою очередь, так называемые «плотные» породы легко рассыпаются в руках, а в воде размокают до глиноподобного состояния. Наглядное отличие этих разновидностей демонстрируют результаты гранулометрического анализа, включающего отмучивание глинистых частиц и рассев более крупного материала (рис.3).
Как видно из приведенных данных, выход гранулометрического класса (–0,01 мм), очевидно, следует рассматривать в качестве показателя активности процессов выветривания и интенсивности развития глинистых минералов по сланцам. В шлифах выветрелые сланцы сложены тонко-микрозернистым агрегатом кварц-иллитового состава. В их строении сохраняются структуры микрокливажа, сланцеватости, а также теневые контуры выщелоченных либо замещенных первичных минералов (рис.4). Состав проб уточняется данными рентгено-дифракционного количественного анализа (см. таблицу).
Минеральный состав глин коры выветривания Тургоякского месторождения, %
|
Разновидности глин |
Минералы и их основные рентгеновские отражения |
||||||||
|
Смектит (12.5-15.5А) |
Хлорит (14.2А) |
Каолинит (7.15А) |
Слюдистые (10.0А) |
Кварц (3.34А) |
Плагиоклаз (3.22А) |
Рутил (1.69А) |
Гетит (4.18А) |
Гематит (2.69А) |
|
|
«Плотные» |
4,0 |
– |
8,1 |
43,8 |
44,1 |
– |
|||
|
Рыхлые |
13,2 |
– |
26,6 |
26,8 |
29,9 |
– |
0,9 |
2,6 |
|
|
17,5 |
– |
16,1 |
30,1 |
15,0 |
21,3 |
– |
– |
– |
|
|
16,1 |
2,7 |
31,8 |
29,0 |
15,4 |
2,8 |
0,6 |
1,6 |
||
Судя по глинистым минералам, рыхлые (диспергированные) глины коры выветривания являются более гипергенно измененными, чем «плотные» разности.
Результаты изучения глин коры выветривания Тургоякского месторождения методами сканирующей электронной микроскопии (СЭМ)
Исследования проводились с использованием прибора Tescan Vega 3, оснащенного рентгеновским спектрометром с энергетической дисперсией INCA x-sight с разрешением 133 эВ и площадью активной зоны кристалла 10 мм2. Сколы образцов размерами не более 2×2 см для лучшей электропроводимости напылялись золотом. Полуколичественный химический анализ выполнялся при ускоряющем напряжении 20 кВ и силе тока 70 нА. Диаметр электронного зонда 0,3-3 мкм, диапазон определения элементов от 0,1 до 100 %. Изучение образцов проводилось в обратнорассеянных (BSE) и вторичных электронах (SE).
Учитывая физическое состояние глин, представления об их строении основываются на изучении «плотных» образцов пород, особенности которых характеризуют более 200 электронно-микроскопических снимков и микрозондовых замеров породообразующих минералов. Анализ собранного материала позволяет утверждать, что в образовании каолиновых глин Тургоякского месторождения активно участвовали разнообразные сообщества микроорганизмов, что иллюстрируют многочисленные снимки фоссилизированных бактериальных структур в виде минерализованных биопленок, псевдоморфоз по колониям и одиночных форм коккоидных и палочковидных бактерий, остатков гликокаликса, нитчатых микрофоссилий, а также микробиоты неясного систематического положения. Распределение микроорганизмов в глинах неравномерное – от единичных остатков до массовых скоплений. Состав минерализованной микробиоты также варьирует в широких пределах – от оксидных до силикатных и алюмосиликатных минеральных разностей.
Характеристика установленной микробиоты
Биопленки и их бесструктурные фрагменты (гликокаликс) являются наиболее распространенными биоморфозами. Они отмечаются по стенкам трещин и пор, на поверхностях минеральных зерен, а также на сколах и по плоскостям сланцеватости выветрелых пород. Предполагается, что подобные образования представляли собой сообщества микроорганизмов, заключенных в слизистое вещество из полисахаридов (экзополисахаридное полимерное вещество, ЭПС), которые вырабатываются бактериальными клетками для защиты от внешних воздействий [31]. В ископаемом состоянии при фоссилизации такого вещества возникают минеральные пленки с бактериальным либо однородным (бесструктурным) строением. С морфологических позиций пленки весьма разнообразные и различаются по внешнему облику, степени раскристаллизации, а также особенностям строения. В последнем случае они могут быть как одно-, так и многослойными с чередованием микропросечек из слюдистых частиц. Поверхности пленок гладкие либо мелкобугристые. Бугристый вид им придают контуры бактериальных клеток, выступающие из-под покрова ЭПС (рис.5, а). Детали внутреннего строения биопленок раскрываются при использовании больших увеличений. Главными структурообразующими формами являются коккоидные и палочковидные бактерии. В многослойных пленках отмеченные микрофоссилии развиваются обособленно друг от друга, образуя в каждом слойке собственную «экологическую нишу».
Палочковидные бактерии – вытянутые клетки цилиндрической формы с закругленными торцами. Палочки прямые, плотно прижатые друг к другу и поэтому имеют близкую пространственную ориентировку, что, вероятно, отражает направление движения поровых вод, необходимых для жизни микроорганизмов. Длина палочек до 25 мкм, толщина – не более 2,5 мкм (рис.5, б). Коккоидные бактерии представляют собой крошечные (0,5-1,0 мкм) шарообразные тельца с четкими, реже расплывчатыми границами. Прилипая друг к другу, они образуют массовые скопления, которые заполняют углубления и обрастают выпуклые поверхности биопленок (рис.5, в). При раскристаллизации биопленок контуры коккоидных клеток, как правило, «стираются», но даже в этих случаях от них сохраняются теневые реликты (рис.5, г). Минерализованные дисковидные бактерии развиваются обособленно от биопленок. Обычно они встречаются группами из 10-15 особей и отличаются от кокков не только своеобразной формой, но и значительно большими размерами, достигая в диаметре 5 мкм при толщине 0,5 мкм. Хорошая сохранность биоты позволяет видеть деление клеток с их последующим превращением в дипло- и тетракокковые формы, а также сарцины (рис.5, д). Аналогичные бактериальные остатки описаны в латеритных бокситах Бразилии и отнесены к цианобактериям рода Synechococcus [23].

Рис.5. Изображения микробиальных структур в каолиновых глинах Тургоякского месторождения во вторичных электронах (SE): а – фрагмент минерализованной биопленки (бп) с признаками замещения агрегата слюдисто-монтмориллонитового состава (мн), хорошо выражено мелкобугристое строение пленки, отражающее контуры бактериальных клеток (бк), заключенных в веществе пленки; б – сообщество плотно прижатых друг к другу палочковидных бактерий (п); в – массовые скопления коккоидных микроорганизмов (к); г – перекристаллизованная биопленка с теневыми реликтами бактериальных кокков (рк) с увеличенным фрагментом на врезке; д – фоссилизированные
Минерализованные микроколонии сложены биоморфозами из сферических клеток, которые, нарастая друг на друга, образуют гроздевидные постройки на стенках трещин (рис.6, а). Для таких клеток характерны четкие границы, одинаковые размеры (8-10 мкм), выраженные следы деления, «морщинистая» поверхность и перекристаллизованное до радиально-лучистых структур внутреннее строение. Часть клеток сфер «сплющены», что может говорить о формировании данной микробиоты сингенетично процессам уплотнения пород.
Биоморфозы нитчатых микрофоссилий представлены остатками цианей, актиномицетов и микромицетов. Чаще всего такая микробиота заселяет участки с отчетливо выраженными признаками разрушения минеральной матрицы породы. При этом фиксируется явная тенденция обособленности друг от друга отмеченных микроорганизмов по видовым «экологическим нишам». Остатки цианей в основном плохой сохранности. Это одиночные прямые или изогнутые сплющенные нитевидные формы, у которых иногда сохраняется ненарушенное строение. В подобных случаях можно видеть фрагменты чехла с сечением 1,5-2,5 мкм и заключенные в нем скрученные в жгуты трихомы (рис.6, б). Минерализованные актиномицеты на электронных снимках напоминают мох. Такое сходство им придает густое переплетение субмикроскопических гифов, образующих пушистый налет на минеральных частицах породы (рис.6, в). Миксомицеты состоят из гифов, слагающих мицелий в виде перепутанных ветвящихся нитей длиной 20-30 мкм. У многих из них на концах наблюдаются утолщения размером 0,5-1 мкм, которые, возможно, являются «плодовыми телами» микрогрибов. Сохранность нитевидных гифов разная: от хорошо выраженных форм до нитей, чьи контуры буквально сливаются с минеральной матрицей породы (рис.6, г).
Биоморфозы микрофоссилий неясного систематического положения характеризуются разнообразной морфологией. Это правильные и сплюснутые шары и эллипсоиды, грибовидные постройки, а также органические остатки в виде чаш и скорлупок. Их размеры варьируют от первых до нескольких десятков микрометров. Поверхность у них гладкая или осложнена наростами из крошечных пластинок и шариков. Внутри биоморфозы могут быть как пустотелыми, так и монолитными. По-видимому, некоторые из них представляют собой остатки минерализованных эукариотов (рис.6, д, е).

Рис.6. Микробиальные структуры в каолиновых глинах Тургоякского месторождения: а – минерализованные гроздевидные микроколонии, в верхнем правом углу их радиально-лучистое внутреннее строение; б – фрагменты остатков чехлов (ч) цианей, внутри которых видны переплетенные в «жгуты» трихомы (т); в – остатки минерализованных актиномицетов (а), как мох обрастающих и замещающих чешуйки слюдистых минералов, с увеличенным фрагментом чешуйки слюды со следами бактериальной коррозии; г – минерализованные гифы (г) миксомицет с утолщениями на концах нитей (плодовые тела? – (пл), в правой и центральной части несколько гифов «сливается» с частицами матрицы породы, что указывает на их одинаковый минеральный состав; д, е – примеры биоморфоз микрофоссилий неясного систематического положения (возможные остатки эукариот?). Изображения а, в, е – во вторичных электронах (SE), б, г – в обратнорассеянных (BSE) электронах; д – в обратнорассеянных и вторичных электронах
Вещественный состав биоморфоз микробиоты
Анализ микрозондовых данных показывает, что минерализованные биогенные остатки сложены широким спектром преимущественно литофильных и сидерофильных элементов. Установлено не менее 20 вариантов различных их сочетаний. Причем химический состав биоты независимо от видовой принадлежности микроорганизмов не является постоянным, а варьирует от оксидных до силикатных и алюмосиликатных минеральных разностей. Несмотря на внешнюю хаотичность в распределении элементов, по характеру их концентраций намечается два типа биоморфоз. Первые по составу близки или аналогичны окружающей минеральной матрице породы (биопленки, коккоидные сообщества, нитчатые организмы), вторые резко от нее отличаются и представляют собой обособленные минеральные фазы (дисковидные бактерии).
Наибольшим разнообразием обладают биопленки. Их состав варьирует от простого оксида кремния до сложных многокомпонентных систем, включающих помимо Al, Fe, Mg, Ca, K, Na примеси Тi, Co, Sb, In, Tb, а также Hf, Sc, Er (рис.7, а). В ряде случаев к такому спектру добавляется углерод, что подтверждает биогенную природу минерализованных пленок. Точный минеральный состав пленок из-за сложного и изменчивого набора слагающих их элементов определить не удается.
Однако необходимые (пусть и косвенные) данные были получены при сопоставлении микрозондовых анализов пленок и минеральных агрегатов, по которым они развиваются. Выполненное сравнение показывает, что в большинстве случаев эти образования химически родственны и, следовательно, в минеральном отношении также должны быть близкими. Таким образом устанавливаются каолинитовый, смектитовый, слюдистый и кремнистый составы пленок. Что касается ранее отмеченных микропримесей, то их наличие, очевидно, указывает на способность биопленок избирательно накапливать элементы, чьи концентрации в минеральной матрице породы ниже чувствительности микрозондового анализа, все это приемлемо и для других видов микробиоты. Однако они явно обеднены микропримесями. В качестве примера на рис.7, б приводятся кремнистые биоморфозы по остаткам микрококковых организмов, которые развиваются на поверхности кварцевого зерна.
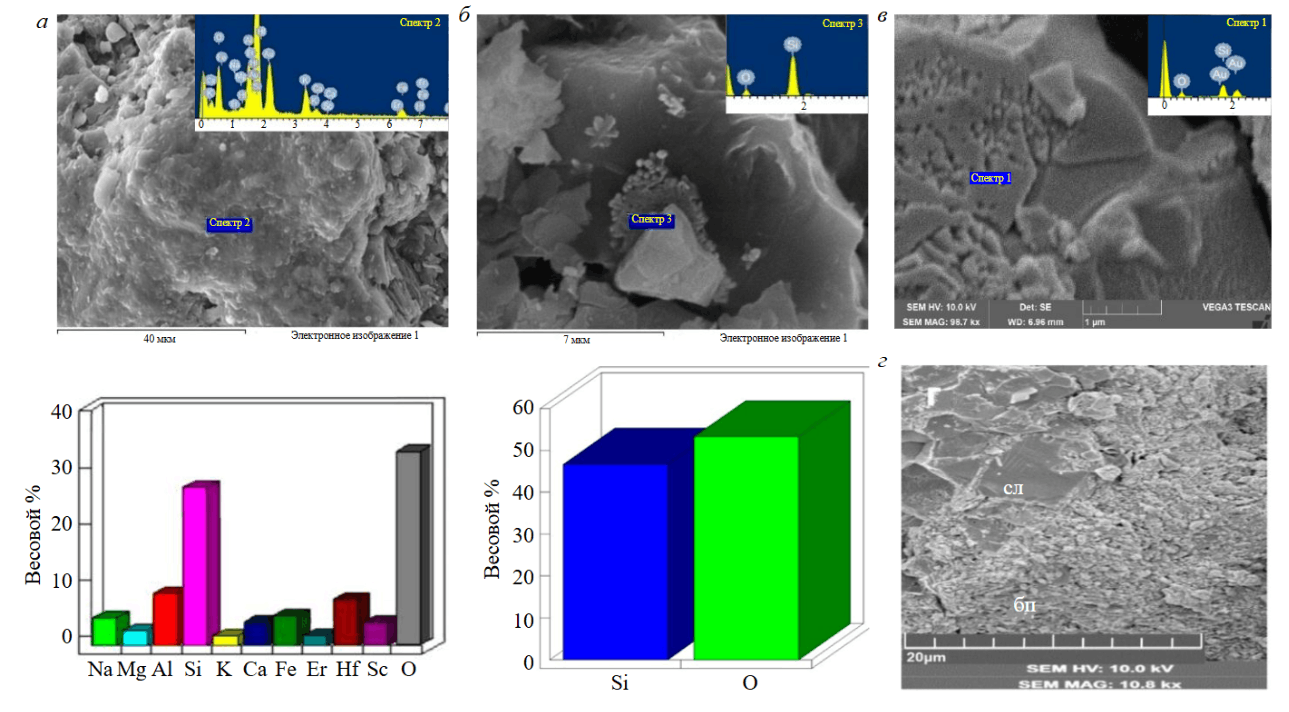
Рис.7. Изображения микробиальных структур в каолиновых глинах Тургоякского месторождения во вторичных электронах (SE): а – фрагмент биопленки (гликокаликс) и ее химический состав (по данным ЭДС); б – биоморфозы по кокковидным клеткам и их химический состав (по данным ЭДС); в – следы бактериальной коррозии зерна кварца; г – замещение слюдистого агрегата (сл) матрицы породы веществом биопленки, хорошо видны реликтовые частицы слюды на поверхности биопленки
Очень важный вопрос – в какой форме находится вещество, слагающее отмеченные биоморфозы? Выполненные наблюдения показывают, что в их строении участвуют как кристаллические, так и аморфные фазы. При этом в ряде случаев (например, для биопленок) фиксируется комбинированное состояние вещества с переходами от бесструктурных рентгеноаморфных (?) к слабо и хорошо раскристаллизованным участкам. Иначе говоря, отчетливо прослеживается трансформация вещества при его перекристаллизации от аморфного до кристаллического состояния. Это важно для понимания механизма образования ранее отмеченных радиально-лучистых структур бактериальных колоний. Очевидно, они также являются результатом раскристаллизации метаколлоидов смектитового минерала.
Иная картина характерна для дисковидных бактерий. Состав их биоморфоз, независимо от окружающих минералов, всегда однотипен и представлен диоксидом титана (см. рис.5, е), иногда обогащенного W и Rb. Здесь явно намечается прижизненная специализация биоты по отношению к титану. Как и в других случаях, точно определить минеральную принадлежность дисковидных биоморфоз не представляется возможным, поскольку известны 11 минеральных модификаций двуокиси титана, из которых наиболее распространенными являются рутил, брукит и анатаз [32].
Роль микробиоты в образовании коры выветривания Тургоякского месторождения
Выявленные морфологические и минералого-вещественные характеристики микробиоты, а также особенности ее взаимоотношений с породообразующими минералами позволяют оценить вклад микроорганизмов в каолинитовое корообразование. Для понимания проблемы целесообразно сопоставить изложенный материал с известными данными о роли микробиоты при формировании латеритных кор выветривания. Работами отечественных и зарубежных ученых установлено повсеместное и разноплановое участие в латеритном выветривании бактериальных сообществ нитчатых, коккоидных и палочковидных организмов, деятельность которых носит как разрушительный, так и созидательный характер [23, 25, 26]. Разрушительный характер выражается в механической деструкции и биохимическом выщелачивании минеральных агрегатов, что приводит к диспергированию выветриваемых пород. Созидательный характер проявляется в аккумуляции растворенного вещества и его отложении в виде псевдоморфоз по биопленкам, остаткам цианей, нитчатым кокко- и палочковидным микрофоссилиям. Возникают метаколлоидные, дисперсные и раскристаллизованные минеральные фазы оксидов и гидроксидов Al, Fe, Mn, Ti.
Сравнительный анализ приведенных материалов по микробиоте латеритов и каолинов Тургоякского месторождения позволяет установить черты сходства и различия биогенных процессов двух типов корообразования. В частности, как и для латеритов, при каолинитовом механизме выветривания активно задействованы одни и те же бактериальные морфотипы, но плотность их заметно меньше, а сами микроорганизмы преимущественно обособляются в разных экологических нишах. Подобно латеритам в каолиновых породах также проявлена разновекторная деятельность микробиоты. С одной стороны, эта деятельность ведет к разрушению минеральной матрицы пород, а с другой – способствует ее восстановлению за счет синтеза новых («биогенных») минералов. Бактериальное разрушение матрицы достигается благодаря механическому и биохимическому воздействию микроорганизмов на минералы. Чаще всего эти два фактора проявляются совокупно, дополняя друг друга. Механический фактор в полной мере проявляется у нитчатых организмов, когда их трихомы или гифы, разрастаясь, проникают между минеральными чешуйками и раздвигают их в разные стороны (см. рис.6, г). Одновременно бактериальные клетки, выделяя метаболиты, химически растворяют окружающие минералы и тем самым подвергают каолиновую породу дополнительному диспергированию. Как показывают наблюдения, следы биохимического растворения можно встретить даже на поверхности кварцевых зерен (рис.7, в).
Бактерии и бактериальные сообщества воздействуют на минералы не только косвенно, но и напрямую, осуществляя их коррозию. В этом случае вдоль контактов возникают структуры замещения (см. рис.6, в) с проникновением биогенной ткани в минеральные зерна, от которых остаются только реликтовые частицы (см. рис.5, а, 7, г). Наблюдается некая связь между биотой и составом замещаемого минерального агрегата. Наиболее активно микробиальной «атаке» подвергаются слюдистые минералы и полевые шпаты, в меньшей степени монтмориллонит и крайне редко – каолинит. Возможным «виновником» такого явления может быть калий. Он, с одной стороны, входит в состав минералов, а с другой – представляет собой химический элемент, необходимый для жизни бактерий. Вероятно, именно по этой причине бактерии в первую очередь корродируют наиболее обогащенные калием минералы, особенно когда он находится в легкодоступном состоянии. Возможное развитие событий установлено в ходе экспериментальных работ и геологических наблюдений [16, 20, 33].
Еще один важный вопрос касается биогенных минералов каолинитовой коры выветривания. Как и в латеритах, они являются важной составляющей формирующихся пород и также представлены псевдоморфозами по биогенным остаткам. Общий механизм их образования разбирается в серии работ российских и зарубежных ученых [31, 34, 35]. Этот процесс подразделяется на минерализацию, биологически индуцированную (BIM) и биологически контролируемую (BCM) [36-38]. В первом случае минералы кристаллизуются на поверхности клеток и их оболочках в ходе реакций продуктов метаболизма с минерализованными поровыми растворами. Во втором случае зарождение и развитие минералов происходит внутри клеток и контролируется внутриклеточными процессами. Несмотря на один и тот же состав биоты и одинаковый механизм образования биогенных минералов, результаты микробной деятельности в условиях латеритного и каолинитового выветривания принципиально различаются. Это наглядно видно на примере составов аутигенных биоминералов – в латеритах они представлены оксидами и гидроксидами Fe, Al, Mn, Ti, а в каолинах это силикатные и алюмосиликатные разности, среди которых резко доминируют глинистые биоминералы коллоидной размерности. При этом в обоих типах кор выветривания состав биоминералов аналогичен минералам абиогенного происхождения, образовавшимся химическим путем в ходе выветривания пород. Отсюда напрашивается вывод, что при формировании коры выветривания, независимо от ее типа, химическое и биогенное минералообразование происходит по одним законам, которые контролируются климатическим фактором и напрямую зависят от состава материнских пород. Как известно, именно эти два показателя определяют активность геохимической среды, ее Еh, pH, температуру, а также интенсивность водного дренажа выветриваемых пород. В свою очередь, геохимические условия выветривания (щелочные для каолинов и кислые для латеритов) определяют способность элементов переходить в растворы и участвовать в химических и биологических реакциях. В латеритах, судя по соотношению хемогенных и биогенных ультрамикроструктур, процессы биогенного минералообразования успешно конкурируют с химическими процессами [24, 25, 39].
В каолинах Тургоякского месторождения деятельность биоты не столь очевидна. Максимально ее роль проявлена в локальных трещиноватых зонах, где биогенные структуры и, следовательно, биоминералы становятся доминирующими в строении породы. Здесь следует обратить внимание на еще один важный момент. В обоих типах рассматриваемых кор выветривания фиксируются остатки микроорганизмов одного и того же облика (кокки, палочки, нитчатые формы), но образуемые ими минералы резко различны – оксиды и гидроксиды в одном случае, силикаты и алюмосиликаты – в другом. Отсюда напрашивается вывод о разном видовом составе микроорганизмов, участвующих в латеритном и каолинитовом корообразовании. Можно заключить, что в каолинитовых корах микробиота, образуя биоминералы, выполняет роль своеобразного промежуточного коллектора, где временно накапливаются растворенные вещества разрушенных выветриванием минеральных агрегатов. В дальнейшем либо происходит консервация биоминералов на неопределенное время, либо при переходе выветривания в латеритную стадию они перерабатываются химическими и микробными процессами в оксиды и гидроксиды Fe, Al, Mn, Ti.
Особое место среди микробиоты занимают «титановые» бактерии, способные аккумулировать ТiO2 (см. рис.5, д). Появление титановых биоморфоз в каолиновых породах противоречит геохимическим особенностям титана, который в гипергенных условиях практически неподвижен. Его концентрации в латеритах и бокситах обычно объясняются накоплением зерен остаточных титановых минералов, которые уцелели после выветривания материнских пород [40-42]. Вместе с тем еще в тридцатых годах прошлого века В.И.Вернадский, опираясь на факты концентрации Тi в организмах, высказывал мысль о существовании в природе бактерий, способных усваивать этот элемент [15], именно их деятельность может быть основной причиной концентрации титана в продуктах конечного выветривания (латеритах и бокситах). Результаты выполненных электронно-микроскопических исследований каолинов Тургоякского месторождения подтверждают правоту гениального ученого.
Заключение
Проведенные исследования показали, что тургоякский каолин представляет собой развитую по сланцам кору выветривания каолинитового типа. В его состав помимо собственно каолинита входят слюдистые и смектитовые минералы, а также в разной степени переработанные частицы полевых шпатов и кварца. Важнейшим элементом строения каолина являются биоморфозы по бактериальным остаткам разной морфологии (кокки, палочки, бактериальные пленки, нитчатые организмы, гликокаликс). В отличие от латеритов их массовое скопление ограничено локальными зонами, особенно в тех местах, где у породы проявлены отчетливые следы разрушенности (пористость, трещиноватость).
Как показывают наблюдения, при создании каолина микробиота участвует в двух процессах – биомеханически разрушает матрицу исходной породы и одновременно с этим синтезируют новые минералы, формируя биоморфозы. Минерализация биоты сопровождается ювелирным копированием морфологических признаков бактериальных клеток, это может быть свидетельством ее принадлежности к биологически индуцированному типу [36, 38]. Принципиальным также является то, что, несмотря на аналогию морфологических типов бактериальных остатков в латеритах и каолинах, состав биоминералов биоморфоз у них совершенно разный – в первом случае это оксиды и гидроксиды Fe, Al, Mn, во втором – силикаты и алюмосиликаты. Это прямо указывает на разный видовой состав микробиоты, участвующей в формировании латеритных и каолинитовых кор выветривания. Отсюда напрашивается еще один вывод – поскольку главным фактором корообразования является климат, то при выветривании он контролирует не только химическое, но и бактериальное минералообразование.
Традиционно скопления титана объясняются концентрациями остаточных титановых минералов, сохранившихся от выветривания материнских пород [40-42]. Однако академик В.И.Вернадский предсказал возможный бактериальный механизм накопления элемента в латеритах и бокситах [15]. В ходе проведенных микрозондовых и электронно-микроскопических исследований каолина Тургоякского месторождения были обнаружены остатки дисковидных бактерий титановой специализации, способных аккумулировать титан, извлекая его из окружающих минералов. Этот факт, с одной стороны, подтверждает точку зрения В.И.Вернадского, а с другой – расширяет наши представления о природе титана в корах выветривания, где он существует не только в авто-хтонных минералах, но и в биогенной форме. Оценка соотношения этих разностей титана является задачей будущих исследований.
Литература
- Полынов Б.Б. Кора выветривания. Ч. 1. Процессы выветривания. Основные фазы и формы коры выветривания и их распределение. Л.: Изд-во Академии наук СССР, 1934. 243 с.
- Петров В.П. Основы учения о древних корах выветривания. М.: Недра, 1967. 343 с.
- Гинзбург И.Н. Древняя кора выветривания на ультраосновных породах Урала. В 2 частях. Часть 2. Геохимия и геология древней коры выветривания на Урале // Труды Института геологических наук. М.: Изд-во Академии наук СССР, 1947. 135 с.
- Слукин А.Д. Коры выветривания и бокситы древних Азиатских платформ: Автореф. дис. … д-ра геол.-минерал. наук. М.: Институт геологии рудных месторождений, петрографии, минералогии и геохимии, 1991. 50 с.
- Добровольский В.В. Гипергенез и коры выветривания. Избранные труды. Т. 1. М.: Научный мир, 2007. 512 с.
- Bowell R.J., Butt C.R.M. Geology, Geochemistry and Formation of Supergene Mineral Deposits in Deeply Weathered Terrain. Springer, 2025. 428 p. DOI: 10.1007/978-3-031-75733-4
- Разумова В.Н., Херасков Н.П., Черняховский А.Г. Геологические типы кор выветривания и примеры их распространения на Южном Урале. М.: Изд-во Академии наук СССР, 1963. 142 с.
- Alcalde-Aparicio S., Vidal-Bardán M., Alonso-Herrero E. Mineralogy and geochemical signatures as indicators of differential weathering in natural soil profiles from the West Asturian-Leonese Zone (NW Iberia) // Earth Sciences Research Journal. 2022. Vol. 26. № 1. P. 55-66. DOI: 10.15446/esrj.v26n1.81087
- Perri F. Chemical weathering of crystalline rocks in contrasting climatic conditions using geochemical proxies: in contrasting climatic conditions using geochemical proxies: an-overview // Palaeogeography, Palaeoclimatology, Palaeoecology. 2020. Vol. 556. № 109873. DOI: 10.1016/j.palaeo.2020.109873
- Мамедов В.И., Макарова М.А., Зайцев В.А. Крупнейшая в мире бокситоносная провинция Фута Джалон-Мандинго (Западная Африка). Статья 3: Влияние геоморфологического фактора и возраста рельефа на распределение, масштабы и качество залежей бокситов // Геология рудных месторождений. 2022. Т. 64. № 5. С. 498-527. DOI: 10.31857/S0016777022050070
- Kai Deng, Shouye Yang, Yulong Guo. A global temperature control of silicate weathering intensity // Nature Communications. 2022. Vol. 13. № 1781. DOI: 10.1038/s41467-022-29415-0
- Gunathilake B.М., Jayawardana D.T., Ratnayake A.S. et al. Gondwana sedimentary rocks of Andigama Basin, Sri Lanka: unraveling weathering dynamics, tectonic setting, and paleoclimate // International Journal of Earth Sciences. 2024. Vol. 113. Iss. 8. P. 2357-2372. DOI: 10.1007/s00531-024-02423-9
- Полынов Б.Б. О геологической роли организмов // Вопросы географии. М.: Государственное изд-во географической литературы, 1953. Сборник 33. Физическая география. C. 45-64.
- Ферсман А.Е. Избранные труды. В 7 т. Т. 5. М.: Изд-во Академии наук СССР, 1959. 858 с.
- Вернадский В.И. Избранные сочинения. В 5 томах. Т. 5. Биосфера I-II. Статьи по биогеохимии. Почвы. Газы. Метеориты и космическая пыль. М.: Изд-во Академии наук СССР, 1960. 422 с.
- Štyriaková I., Štyriak I., Nandakumar M.P., Mattiasson B. Bacterial destruction of mica during bioleaching of kaolin and quartz sandsby Bacillus cereus // World Journal of Microbiology and Biotechnology. 2003. Vol. 19. Iss. 6. P. 583-590. DOI: 10.1023/A:1025176210705
- Sedlakova-Kadukova J., Marcincakova R., Luptakova A. et al. Comparison of three different bioleaching systems for Li recovery from lepidolite // Scientific Reports. 2020. Vol. 10. № 14594. DOI: 10.1038/s41598-020-71596-5
- Schmitz A.M., Pian B., Medinet S. et al. Generation of a Gluconobacter oxydans knockout collection for improved extraction of rare earth elements // Nature Communications. 2021. Vol. 12. № 6693. DOI: 10.1038/s41467-021-27047-4
- Авакян З.А., Каравайко Г.И., Мельникова Е.О. и др. Роль микроскопических грибов в процессах выветривания пород и минералов пегматитового месторождения // Микробиология. 1981. Т. 50. Вып. 1. С. 156-162.
- Каравайко Г.И. Микробная деструкция силикатных минералов // Труды Института микробиологии им. С.Н.Виноградского: Юбилейный сборник к 70-летию института. Вып. XII. М.: Наука, 2004. С. 172-196.
- Song W., Ogawa N., Oguchi C.T. et al. Effect of Bacillus subtilis on granite weathering: A laboratory experiment // Catena. 2007. Vol. 70. Iss. 3. P. 275-281. DOI: 10.1016/j.catena.2006.09.003
- Антошкина А.И. Бактериальное породообразование – реальность современных методов исследований // Ученые записки Казанского университета. Серия: Естественные науки. 2011. Т. 153. Кн. 4. С. 114-126.
- Школьник Э.Л., Жегалло Е.А., Богатырев Б.А. и др. Биоморфные структуры в бокситах (по результатам электронно-микроскопического изучения). М.: Эслан, 2004. 112 с.
- Слукин А.Д., Боева Н.М., Жегалло Е.А., Зайцева Л.В. Биоминералы латеритных бокситов – новые данные по результатам электронно-микроскопического изучения // Новые данные о минералах. 2016. Вып. 51. С. 52-61.
- Новиков В.М., Бортников Н.С., Боева Н.М. и др. Биогенные наноминералы оксидов железа в корах выветривания базальтов континентальных окраин Восточной Азии на примере Дальнего Востока России и Вьетнама. Статья 3. Магнетит // Вестник Воронежского государственного университета. Серия: Геология. 2017. № 2. С. 69-73.
- Xiaoxue Yang, Yanzhang Li, Yan Li et al. Microbially induced clay weathering: Smectite-to-kaolinite transformation // American Mineralogist. 2023. Vol. 108. № 10. P. 1940-1947. DOI: 10.2138/am-2022-8442
- Kawano M., Tomita K. Microbiotic Formation of Silicate Minerals in the Weathering Environment of a Pyroclastic Deposit // Clays and Clay Minerals. 2002. Vol. 50. Iss. 1. P. 99-110. DOI: 10.1346/000986002761002865
- Наймарк Е.Б., Ерощев-Шак В.А., Чижикова Н.П., Компанцева Е.И. Взаимодействие глинистых минералов с микроорганизмами: обзор экспериментальных данных // Журнал общей биологии. 2009. Т. 70. № 2. С. 155-167.
- Переломов Л.В. Роль взаимодействий бактерий и глинистых минералов в педохимических процессах // Геохимия. 2023. T. 68. № 10. С. 1021-1031. DOI: 10.31857/S0016752523100102
- Соколова Т.А. Роль почвенной биоты в процессах выветривания минералов (обзор литературы) // Почвоведение. 2011. № 1. С. 64-81.
- Астафьева М.М., Герасименко Л.М., Гептнер А.Р. и др. Ископаемые бактерии и другие микроорганизмы в земных породах и астроматериалах. М.: Палеонтологический институт им. А.А.Борисяка РАН, 2011. 172 с.
- Тхант Зин Пью. Разработка направленного метода получения фотоактивных неорганических покрытий на основе диоксида титана, модифицированного медью: Автореф. дис. … канд. хим. наук. М.: Российский химико-технологический университет имени Д.И.Менделеева, 2023. 16 с.
- Štyriaková I., Štyriak I., Oberhänsli H. Rock weathering by indigenous heterotrophic bacteria of Bacillus spp. at different temperature: a laboratory experiment // Mineralogy and Petrology. 2012. Vol. 105. Iss. 3-4. P. 135-144. DOI: 10.1007/s00710-012-0201-2
- Тихомирова Н.С., Орлеанский В.К. Моделирование фосфатоосаждения в лабораторных культурах цианобактерий // Литология и полезные ископаемые. 1994. № 1. С. 135-140.
- Weiner S., Dove P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect // Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. № 1. P. 1-29. DOI: 10.2113/0540001
- Lowenstam H.A., Weiner S. On Biomineralization. Oxford University Press, 1989. 333 p. DOI: 10.1093/oso/9780195049770.001.0001
- Bazylinski D.A., Frankel R.B. Biologically Controlled Mineralization in Prokaryotes // Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. № 1. P. 217-247. DOI: 10.2113/0540217
- Frankel R.B., Bazylinski D.A. Biologically Induced Mineralization by Bacteria // Reviews in Mineralogy and Geochemistry. 2003. Vol. 54. № 1. P. 95-114. DOI: 10.2113/0540095
- Xiluo Hao, Kwunlun Leung, Rucheng Wang et al. The geomicrobiology of bauxite deposits // Geoscience Frontiers. 2010. Vol. 1. Iss. 1. P. 81-89. DOI: 10.1016/j.gsf.2010.06.001
- Юдович Я.Э., Кетрис М.П., Рыбина Н.В. Геохимия титана. Сыктывкар: Геопринт, 2018. 431 с.
- Страхов Н.М. О значении титанового модуля для познания генезиса бокситов // Литология и полезные ископаемые. 1963. № 2. С. 249-252.
- Бушинский Г.И. Геология бокситов. М.: Недра, 1975. 411 с.